
空气温度对植物的影响
——CANNA研究中心
不足为奇的是,人们已经开展了大量的工作,来研究有效的温室栽培,并提供适宜的温度。然而,植物的最适宜温度取决于一系列因素。植物对周围大气温度的反应取决于它所处的生长阶段。植物体内有生物钟,决定着它对温度的敏感性。
空气温度和植物温度之间的差异
温度越高,大多数生物过程发生速度越快,这一点既有积极影响,也有负面影响。例如,在大多数情况下,可以带来更快的生长速度,提高水果作物的产量。然而,呼吸作用发生会带来负面影响,因为它意味着为果实发育提供的能量更少,果实会更小。有些影响是短期的,有些则是长期的。例如,植物的同化平衡受到温度的影响是即时的,而花朵的形成则是由气候环境在更长的时间内决定的。
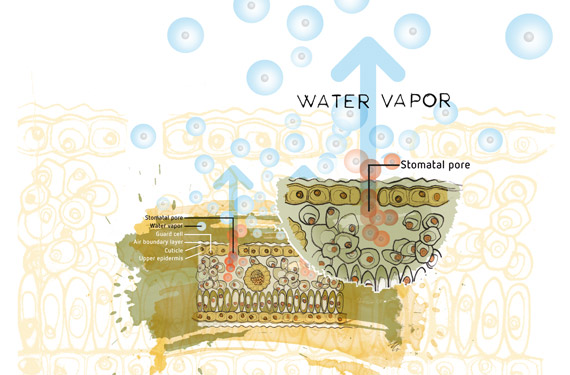
我们可以用高速公路上的汽车来打个比方。气孔是允许汽车离开高速公路的出入口。当有很多汽车堵在出入口时,汽车就不得不放慢速度,于是便发生了交通堵塞。汽车少了,交通流速就会加快。同样的事情也发生在空气分子和空气中的水蒸气分子上。如果在气孔(出入口)周围有更高浓度的分子,那么它们离开气孔的速度就会慢一些,而且会出现堵塞。这就是VPD高时的情况。这就意味着植物的冷却能力较差,会引发植物压力。此外,水分会凝结在叶子表面,形成一层薄膜,为病原体的繁殖提供完美环境。
植物的温度和空气的温度并不相同,因为植物能够通过蒸发降温,在日光照射下升温。植物寻求自身最佳温度,而空气温度、相对湿度和光照之间能达到平衡是极为重要的。如果光照水平高,植物会升温,导致植物温度和空气温度之间出现温差。为了降温,植物就会增加蒸腾速率。除了温度,蒸腾速率还取决于环境条件,如光照、大气二氧化碳水平和相对湿度等,但也与植物的种类有关。
植物由不同的部分组成,不同部分对温度的反应各不相同。果实的温度与空气的温度基本一致;当气温升高时,果实温度也随之升高,反之亦然。但果实的温度波动要小于气温,而且比气温的上升或下降需要更长的时间(有时多出几个小时)。相比之下,花的温度要高于气温或叶片温度,花瓣的蒸腾速率远低于叶片。冠层顶部的温度会比冠层底部的温度产生更大的波动。顶部也更易受日光照射而升温,因此当光照强烈时,顶部会达到比气温更高的温度。
蒸汽压差
环境的相对湿度取决于温度和风速。较高的温度通常会导致蒸腾作用增强。这不仅因为分子移动速度变快,且温暖的空气可以容纳更多的水蒸气。当无风时,叶子周围的空气就会变得水蒸气饱和,从而使蒸发过程减缓。如果空气中充满了水汽,叶片和叶片周围就会凝结一层水膜,这会为病原体提供良好的繁殖环境,进而攻击植物。

蒸汽压力缺口(VPD)可以比作汽车上的转速表。当发动机转速增加时,转速计数器上的指针转向并进入红色区域。这不会立即损坏电机,但如果汽车继续这样驾驶较长时间,就会发生损坏。同样的情况也适用于植物:当VPD在较长一段时间内维持过高水平时,植物无法在夜间恢复,就会发生不可逆转的植物损伤(例如叶片或者花瓣烧枯)。
蒸汽压力缺口(VPD)指的是空气和饱和点之间的水汽含量差。VPD越高,植物通过蒸腾释放的水分越多。然而,如果VPD过大,植物就会受到压力,无法补充通过蒸腾失去的水分。这在短时间内不会造成问题,植物会在之后的夜晚吸收足够的水分来恢复。但是若VPD长时间保持在高水平时,植物无法在夜间恢复,就会发生不可逆转的损伤,例如叶片或者花瓣烧枯。
测量叶片厚度可以从视觉上预测植物的恢复潜力。事实上,叶子在白天会变得更薄,因为它们在蒸腾作用下失去了水分,但如果叶子在某一晚比前一晚更薄,就表明植物未能恢复。因此,似乎保持较低的VPD水平可以避免任何损害。但在这样的条件下,无法充分刺激植物生长,活跃度也不高,当植物面临应激情况时,可能会产生负面后果。
总的来说,这可以与汽车的转速表进行类比。随着发动机转速的增加,转速表的指针上升,进入红色区域。这不会立即损坏引擎,但如果指针在红色区域停时间留过长,则会造成损害。对于大多数植物来说,VPD应该保持在0.45到1.25之间,单位为千帕(kPa,压力单位),最佳值为0.85kPa。VPD与日光照射强度大致相同。在早晨,当阳光开始照射时,VPD上升,在中午左右达到顶峰,随后又逐渐下降。要计算VPD,必须先知道空气温度、植物温度和相对湿度。

大气中的大部分水是以水蒸气的形式存在的。水蒸气虽然不可见,但我们可以通过人体舒适程度来注意到它的存在(较高的湿度会让我们感到粘乎乎的不舒服)。空气能见度也受到水蒸气含量的影响。云是可见的,因为云中包含的水蒸气已经冷却到一定程度,水分子开始凝结,在空气中形成微小的水滴甚至冰晶。因此肉眼可以看见云。
气孔
植物能够调节蒸腾和冷却的过程,借助的特殊植物器官称为气孔。气孔是叶子上的小洞,可以张开或闭合,从而限制水蒸气的进出量。温度越高,气孔打开程度越大。气孔打开程度的测量较为困难,可以利用VPD来进行估算。随着气孔打开程度越来越大,更多的气体可以进出叶子。
环境因素会影响气孔张开程度(导度)——例如,较高的相对湿度会使气孔导度更高,而较高的二氧化碳水平会降低气孔导度。气孔导度也受到环境因素之外的其他因素影响,比如植物激素和植物接收的光的颜色(波长)。植物激素脱落酸会调节气孔中的离子浓度,使气孔在几分钟内迅速打开。波长较短的光(大约400-500纳米,也就是蓝光)比波长较长的光(大约700纳米,也就是红光)使气孔张开得更大。
最佳昼夜温度
植物在白天和夜间会发生不同的生理过程,因此最适宜的昼夜温度也有所不同。糖的运输主要在夜间进行,主要是向植物较温暖的部位运输。叶子比果实和花朵冷却得快,因此大部分可用的能量都进入了植物的果实和花朵,以满足生长发育所需。
1949年,在加州理工学院的一间温室里,研究人员在世界上第一个有空调的房间里研究了植物的最佳昼夜温度组合。实验表明,在光照期高温和黑暗期低温相结合的条件下,番茄植株高度要高于温度不变环境下。植物区分昼夜温度变化的能力被称为温周感应,它对开花、结果和生长都有影响。
当夜间温度升高时,运输到正在生长的组织中糖的数量(在这些组织中,需要能量来支持更高水平的呼吸作用)会受到限制,因此生长也会受到限制。研究还发现,白天高温和夜间低温相组合的条件下,茎会伸长。夜间较低的温度可以改善植物体内的水分平衡,这是茎伸长增加的主要原因。因此,温度可以作为调节植物高度的工具,但夜间较低的温度也可以节省能源。温度对植物形态的周期性影响可以用专业术语"热形态建成"来表示。

最佳空气温度还取决于光照强度和空气中二氧化碳的含量。植物的生理功能与冷血动物相似,它们的新陈代谢和光合作用的速率随环境空气温度的升高而增加。当温度极低时(具体值取决于植物的种类),无论光照量如何,几乎都不会发生任何光合作用。随着空气温度的升高,光合作用的速率也会增加。当光线和温度达到平衡时,环境中的二氧化碳水平将成为限制因素。如果有足够的二氧化碳,光合作用的速率将随着温度的升高而增加,此外,其他因素也发挥了作用,如RuBisCo酶。
RuBisCo对光合作用来说至关重要。在某些情况下,植物会进行一种称为光呼吸作用的过程,也就是RuBisCo与氧气结合,而不是像正常光合作用中与二氧化碳结合。低光照强度下,二氧化碳含量水平和最佳温度范围都比高强度光照下低,而酶的活性在较高温度下也会增加。
昼夜温差(DIF)
DIF这一概念关注的是昼夜温度之间的关系。日间温度变化对植物茎纵向伸长的影响,主要取决于昼夜温差(DIF),即白天温度减去夜间温度,而不仅仅是单一的白昼或夜间温度。换句话说,温差,以及白昼还是夜晚的温度更高,才是最重要的。
DIF对叶片的生长影响不大,但对节间茎的生长影响较大。正DIF条件下,植株高度要大于零DIF条件下的植株高度,零DIF条件下的植株高度要大于负DIF条件下的植株高度,且节间更长。负DIF(即白天温度低于夜间温度)还会对植物产生其他重要形态发育影响,包括叶柄、花茎、花梗和叶的长度更短。

节间伸长以及叶片伸长出现差异,是由细胞伸长以及细胞分裂过程的差异造成的。当DIF为负时,这两个过程都受到抑制,这可能是由于根尖的近顶端分生组织(一种负责生长的植物组织)中赤霉素活性有所降低。赤霉素是一种刺激植物生长的植物激素。在快速生长期,DIF对茎伸长的影响最大,幼苗对昼夜温差的敏感性要高于成株。因此,保持DIF为负,对于茎伸长早期限制植株高度来说尤为重要。
茎的伸长也可能是由于24小时的生长周期中温度快速下降(约两个小时)造成的,这通常出现在日光刚刚出现之时或者之前,以及夜间。无论是长日植物、短日植物,还是中日植物,对温度变化的反应在白昼光照期的前几个小时最为强烈。因此,夜间最后两个小时,气温的下降会对植株高度产生影响。在凉爽地区的秋季,由于夜间自然温度较低,上述环境很容易在温室内实现。
茎伸长在日间和夜间对温度敏感性的变化可能受内源生长节律的控制。1994年,人们发现菊花的生长具有24小时左右的昼夜节律。植物茎的伸长在24小时的昼夜循环中并非恒定。在诱导开花条件下,短日植物和长日植物在夜间比白天伸长得更快。而兰花则需要经历夜间低温期才能开花。
种植者经常会采用一种叫做积温的策略,确定作物生长的最低和最高温度,允许温度在较长时期内保持平均温度的前提下,存在一定变化。这一策略最大程度上利用了自然的热量。
气温是影响植物生长发育和生长速度的主要环境因素。然而气温从来都不是一个孤立的因素。植物生长中的每一个因素都与其他因素息息相关,种植者面临的挑战就在于找到其中的薄弱环节。本文探讨了多个因素,但此外还有其他同样重要的影响因素,比如水分平衡,以及蒸腾作用等等。植物的一切活动都是在以气温为前提下进行的,正确处理气温问题,是成功栽培作物这一漫长道路上的第一步。


